Osmóza aneb jaké to je být rybou ve vodě
Osmóza je fyzikální jev, který zásadním způsobem ovlivňuje životní procesy a zdraví našich ryb. Přesto plný význam slova málokdo zná, ačkoliv každý akvarista dnes ví, že existuje tzv. reverzní osmóza. Nebudeme se ale prozatím věnovat technice. Pojďme si zodpovědět otázky, jestli ryby pijí, proč je zrovna pro vývoj jiker často tak důležitá měkká voda a proč (a jestli vůbec) rybám pomáhá, když vodu „přisolíme“.
Základní pojmy
Difúze – Fyzikální jev, při kterém dochází k pohybu částic z prostředí s vyšší koncentrací do prostředí s nižší koncentrací. Klasickým příkladem je rozpouštění soli ve vodě – není potřeba míchat, po určitém čase bude voda v celém svém objemu stejně slaná. Difúze je jev pasivní, tzn. k pohybu částic není zapotřebí vynaložit žádnou energii.
Osmóza – Speciální případ difúze. Máme dva roztoky s rozdílnou koncentrací, oddělené polopropustnou membránou (tzn. částice menších rozměrů mohou skrz membránu procházet). Přes membránu v tomto případě prochází rozpouštědlo, v živých systémech zpravidla voda. Osmóza tedy rovněž směřuje k tomu, že voda bude všude stejně „slaná“, ale za tu cenu, že voda přeteče do původně „slanějšího“ roztoku tak, aby ho naředila a koncentrace na obou stranách membrány se vyrovnaly.
(Pozn. Pokud se nekamarádíte s fyzikou a chemií a text v podobném duchu by vás odradil od dalšího čtení, přeskočte na další nadpis níže. Není možná až tak zásadní vědět, PROČ a JAK se našim rybám něco děje. Ale děje se a já budu ráda, pokud si z článku odnesete alespoň poznání, CO a jaké to má důsledky.)
Osmotický tlak – Jak vyplývá z výše uvedeného, osmotická síla vedoucí k vyrovnání koncentrace může způsobit, že objem roztoků oddělených membránou bude ve výsledku odlišný – a osmotická síla působí dokonce proti síle gravitační, jak je vidět na ilustraci. Tzv. osmotický tlak odpovídá tlaku, který by bylo potřeba vyvinout, aby se hladiny vyrovnaly (nebo si můžeme představit tlak, vyvíjený na „slanější“ roztok, který by zabránil přitékání vody přes membránu; vyvineme-li tlak ještě větší a donutíme vodu téct na opačnou stranu, voilà, máme reverzní osmózu). Tato teoretická definice nás nemusí mást – podstatné je si představit, že pokud jsou dva různě ředěné roztoky oddělené membránou, nějaký osmotický tlak existuje = dochází k toku rozpouštědla přes membránu. Pozor, osmotický tlak nezávisí jen na prosté koncentraci látek.
Osmolarita, osmolalita – Celková látková koncentrace osmoticky aktivních částic. Udává se v Osm/l (osmolarita, také Osmol/l nebo mol/l) nebo Osm/kg (osmolalita, i tady najdeme varianty Osmol/kg či mol/kg). Pro nás jde o pojmy ne zrovna zásadní, jen je potřeba si uvědomit, že pokud se bavíme o koncentraci roztoků na obou stranách membrány, ne vždy srovnáváme chemicky totožné látky. Krev, cytoplazma atd. nejsou tvořeny stejnými látkami, přesto mohou mít stejnou osmolalitu jako čistá voda se špetkou kuchyňské soli.
Osmotická rovnováha – Stav, kdy osmolalita na obou stranách membrány je stejná.
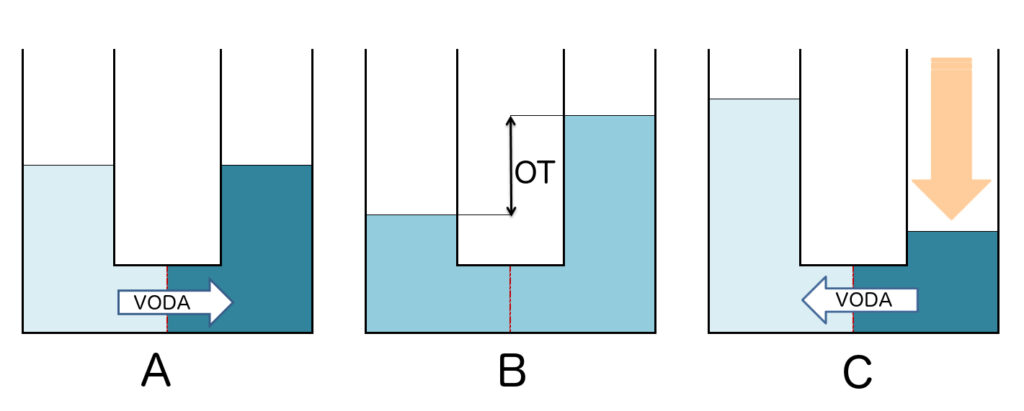
Obr. A: Výchozí stav, voda protéká skrz membránu do roztoku s větší osmolalitou.
Obr. B: Osmotická rovnováha. OT = osmotický tlak, v jehož důsledku došlo k vyrovnání osmolality obou roztoků, ale změnil se jejich objem.
Obr. C: Reverzní osmóza vede k proudění vody opačným směrem proti působení osmotického tlaku jako následek silnějšího tlaku z vnějšího zdroje (oranžová šipka).
Co je podstatné, k difúzi i osmóze v případě nerovnováhy prostě dochází – nelze je zastavit. Pro fungování všech živých organismů má osmóza obrovský význam. Obsah buněk a tím i orgánů a celého těla je od okolí oddělen polopropustnou membránou, takže buňka a potažmo i celý organismus jsou s tímto pohybem vody dovnitř nebo ven neustále konfrontovány. Že může jít o skutečně masivní dopad, ilustruje jeden názorný příklad:

Rybička nasála tolik vody, že je její struktura velmi narušená a sotva drží pohromadě. Pokud vám to připomíná skutečnou rybu s vodnatelností, není to náhoda, ale nepředbíhejme. Přijímání vody je každopádně jen jednou možnou reakcí, pokud je vnitřní prostředí potřeba naředit. Pokud bychom rybičku či jiný vhodný objekt naopak namočili do velmi slaného nálevu, došlo by ke ztrátě vody a svraštění. Zkuste posolit rozkrojenou cibuli, jak se orosí – důvodem je opět osmóza.
Vnější prostředí může být ve vztahu k vnitřnímu (např. buňce) podle koncentrace rozpuštěných solí: Isotonické = má stejný obsah soli. Hypertonické = má vyšší obsah soli, což vede k tendenci buňky ztrácet vodu a ředit tím okolní roztok.Hypotonické = má nižší obsah soli, tj. buňka naopak vodu přijímá, čímž ředí svůj obsah.
Existuje sice pojem isoosmotický (příp. hyperosmotický, hypoosmotický), ale pro zjednodušení si vystačíme s tonicitou. Ta se vztahuje k osmotickému tlaku. Některé látky s velmi drobnými částicemi mohou membránou také procházet, takže i když má roztok stejnou osmolalitu (= stejný počet částic na obou stranách membrány), nemusí být isotonický, protože zde působí určitý osmotický tlak. To je ale pro naše porozumění už příliš komplikované a nadbytečné. Podstatné je, že je-li roztok isotonický, tlak je vyrovnaný a voda přes membránu jednosměrně neproudí.
Jak vlastně souvisí osmóza a obsah solí? Nemyslím samozřejmě jen NaCl. Osmoticky aktivní jsou obecně látky rozpuštěné ve vodě, anorganické i organické – u živých organismů především soli, cukry, bílkoviny. Soli hrají za běžných podmínek nejvýraznější roli (protože jsou disociované na anionty a kationty, tzn. jde o elektrolyty, proto je jejich tonicita velká) a NaCl je ideální látkou, s pomocí které lze osmózu popisovat, modelovat a především ovlivňovat.
Nasnadě je otázka: když buňka vodu přijímá/ztrácí, kdy se proces zastaví? Pokud nedojde k vyrovnání koncentrací, nezastaví se. Buňka praskne, nebo dojde k jejímu nevratnému poškození ztrátou vody. Daleko odolnější jsou rostlinné buňky, které jsou chráněny pevnou buněčnou stěnou, tedy neprasknou; při nedostatku vody u nich dojde k odloučení cytoplazmatické membrány obklopující obsah buňky od stěny, smrskává se tedy jen „vnitřek“ a proces je vratný. U živočišné buňky dochází k prasknutí přetlakem nasáté vody velmi snadno; při ztrátě vody se buňka celá svrašťuje, a pokud tento stav trvá delší dobu, je nevratný.
Tolik fyzikální pozadí tohoto všudypřítomného jevu. Předchozí odstavec naznačil, že osmóza může vést k závažnému poškození organismu. Proto bychom měli vědět, jak na ryby působí a na co si dávat pozor. Úplně posledním strašákem na úvod budiž osmotický šok, tedy stav, kdy dojde k náhlé a výrazné změně osmotického tlaku. Přelovíme-li sladkovodní rybu do mořského akvária, bude to právě osmotický šok, který jí způsobí vážné zdravotní komplikace.
Jak se živočichové s osmózou vyrovnávají
Většina obratlovců se snaží o jedno – mít v rovnováze mimobuněčné a vnitrobuněčné prostředí. Tím drží buňky v jakési ochranné lázni a nemusí řešit na této nejnižší úrovni neustálý tok vody dovnitř či ven. Proto obsah našich buněk, krevní plazma, ale např. i slzy jsou navzájem isotonické. Zásadní komplikace ovšem nastávají, pokud je živočich bez této ochrany (tj. primitivní organismy jako jsou prvoci, bakterie apod.) a/nebo pokud žije ve vodním prostředí.
Osmokonformer – Tento organismus udržuje své vnitřní prostředí v rovnováze s vnějším, resp. aktivně nereguluje svou osmolalitu; je to strategie mořských bezobratlých, pro které se dá říci „jaká voda, takové tělo“ (vodní prostředí je isotonické). Ve sladké vodě, která má velmi nízkou osmolalitu (většinou do 30–50 mOsm/l), by nebylo možné takto přežít.
Osmoregulátor – Nepřizpůsobuje se okolí, ale udržuje stabilní množství rozpuštěných látek ve svém těle a s hypotonickým (sladké vody) či hypertonickým (moře) okolním prostředím se vyrovnává různými mechanismy.
Protože jsou nižší živočichové daleko více vystaveni přímému vlivu vodního prostředí, jsou citlivější na změny. Snáze u nich dochází k osmotickému šoku. To je také důvod, proč na vnější parazity může zabírat solná koupel (u sladkovodních ryb), příp. krátkodobé přelovení do sladké vody (u ryb mořských). Zatímco ryba má své regulační mechanismy, které přinejmenším oddálí důsledky nehostinných podmínek, prvok na povrchu jejího těla je hypo- či hypertonickému prostředí vystaven náhle a bezprostředně. Jeho buňka praskne nadbytkem nasávané vody, nebo naopak vodu ztratí a zahubí ji dehydratace. Toho se mj. využívá i při konzervaci potravin – slaný či sladký nálev dehydratací zničí bakterie a další organismy. Osmóza je mocná čarodějka.
Bezobratlí (s výjimkou mořských) využívají různé osmoregulační mechanismy v závislosti na typu vylučovací soustavy. Aktivně pracují s vylučováním či příjímáním vody a solí.U starobylých obratlovců najdeme dva zvláštní typy strategií, jak s osmózou naložit. Sliznatky, což je nejprimitivnější skupina dnes žijících obratlovců, udržují rovnováhu s mořskou vodou. Ta obsahuje asi 1000 mOsm/l rozpuštěných anorganických solí, vnitřní prostředí sliznatek zhruba stejné množství. Jsou to tedy klasičtí osmokonformeři, jediní mezi obratlovci.
Příčnoústí, skupina paryb, kam patří žraloci a rejnoci, jsou někde na půl cesty mezi nejprimitivnějším a nejmodernějším přístupem a využívají k tomu unikátní mechanismus. Všichni obratlovci (s výjimkou výše uvedených sliznatek), včetně těch suchozemských, mají vnitřní obsah anorganických solí přibližně 250–450 mOsm/l. Příčnoústí taktéž, ale aby se vyrovnali mořské vodě, spoléhají na organické sloučeniny. Jejich tělní tekutiny obsahují značné množství močoviny. Tím se dostávají na celkový obsah solí těsně nad 1000 mOsm/l. Prakticky tak eliminují ztráty vody v jinak silně hypertonickém prostředí, ale musí se pro změnu vyrovnat s toxicitou močoviny (k tomu významně pomáhá trimethylaminoxid, jehož hladina v krvi je rovněž vysoká) a s nadbytkem soli, kterou vylučují speciální rektální žlázou.Žraloci a rejnoci potřebují vysokoproteinovou stravu, aby mohli neustále produkovat dostatek močoviny. Stejně to funguje i u sladkovodních druhů, ale ty udržují obsah organických látek v tělních tekutinách mnohem nižší.
Mořské ryby udržují svou vnitřní iontovou rovnováhu na úrovni kolem 400 mOsm/l. To znamená, že okolní mořská voda obsahuje téměř třikrát více solí. V důsledku toho mořské ryby neustále ztrácejí vodu, která uniká z jejich těla ve snaze o zředění oceánu… Mořská ryba dělá to samé, co děláme my, abychom nahradili vodu ztracenou pocením, dýcháním a nezbytnými fyziologickými procesy – pije. Polyká vodu, která je vstřebána střevní stěnou. Ta je samozřejmě prostupná pro větší částice, zde neprobíhá osmóza. Voda se tak do těla dostává s velkým množstvím solí, což je pochopitelně nežádoucí. Sůl je aktivně vylučována speciálními buňkami v žábrách (především Na+ a Cl–), částečně v koncentrované moči (Mg2+ a SO42-), vápník ve střevě. Mořské ryby tedy v důsledku osmózy musí vynakládat energii na to, aby se zbavily přebytečné soli. Neustále pijí a vylučují jen minimální množství moči (denně do 1 % jejich tělesné hmotnosti).
Sladkovodní ryby, jak už je jistě zřejmé, mají opačný problém. Jejich těla jsou řádově „slanější“ (obvykle zhruba 250–350 mOsm/l) než okolní voda. Ta proto ve velkém množství proniká dovnitř. Částečně celým povrchem těla, ale to je chráněno šupinami a slizovým obalem, z valné většiny tedy dochází k osmóze skrz žaberní lamely, které pochopitelně musí být z důvodu své funkce nejprostupnější. Sladkovodní ryby vůbec nepijí, zato produkují velké množství moči, odpovídající denně 20–30 % jejich tělesné hmotnosti.
Ačkoliv je moč velmi ředěná, přesto při jejím vylučování dochází k určité ztrátě solí a tím i životně důležitých prvků. Ty se navíc ztrácejí přes žábry difúzí. A znovu obrácená situace oproti moři – sůl zde není snadné získat. Ryby musí investovat energii, aby soli aktivně přijímaly. Slouží k tomu speciální buněčné pumpy na žaberních obloucích, kdy za využití energie dochází k přenosu kationtu sodíku přes membránu výměnou za vodík, zatímco chlorid je získáván výměnou za hydrogenuhličitan. Dalším zdrojem je potrava.
Ačkoliv NaCl tvoří jen zlomek obsahu solí ve sladké vodě (na rozdíl od mořské, kde představuje 85–99 % salinity), pro vnitřní prostředí rybího organismu jsou sodík i chlorid velmi podstatné a tvoří asi polovinu anorganických solí. Dalšími klíčovými elektrolyty, nezbytnými pro fungování organismu, jsou Ca2+, Mg2+, K+, PO43-. Ale význam elektrolytů je téma pro samostatný článek, teď se vraťme k osmóze konstatováním, že pro její správné „zvládání“ je nezbytná dostupnost různých prvků, zejména vápníku. Koncentrace elektrolytů je udržována stabilní jejich vázáním v nedostupných formách a opětovným uvolňováním. Vzhledem k neustálé výměně s okolní vodou je potřeba zajistit vnější zdroj pro doplňování ztrát. I to je jeden z důvodů, proč extrémně měkká voda není pro chov ryb vyhovující.
Ledviny sladkovodních ryb jsou poměrně velkým párovým orgánem a vykonávají neustále značné množství práce. Pokud nejsou plně funkční, ať už z důvodu nějaké choroby či oslabení (otrava, infekční nemoc, nevhodná výživa, stres atd.), může dojít k hromadění vody v těle. To pozorujeme jako vodnatelnost. Prvotní příčina může být tedy různá, bezprostřední příčinou je selhávání ledvin – a pomoc je velmi obtížná až nemožná. Může pomoci přisolit vodu v akváriu, a to nejčastěji buď kuchyňskou solí NaCl, nebo tzv. epsomskou (hořkou) solí, MgSO4*7H2O. Eventuelně můžeme použít mořskou sůl, která je vhodná proto, že mimo NaCl dodává i spoustu dalších mikroprvků. Přisolením snížíme rozdíl mezi iontovým obsahem uvnitř a vně rybího těla, takže bude méně vody vnikat dovnitř. To může vést k úlevě, ryba nepotřebuje tolik energie, aby s osmózou bojovala, navíc tím chráníme vnitřní orgány před poškozením „převodněním“. Prvotní příčinu to ale neodstraní, i když vítaným podpůrným efektem může být už zmíněné zahubení případných vnějších parazitů.
Mimo selhání ledvin může k nadměrnému zavodňování vést také poškození povrchu těla či ochranného slizového obalu. Pokud je většího rozsahu, tak kromě zvýšeného vystavení patogenům se stává postižené místo také prostupnější pro vodu. I když je funkce ledvin zachována, do těla se dostává více vody, než je obvyklé a zvládnutelné.
Není nijak překvapivé, že prevence je klíčem ke zdraví: čistá voda se správnou teplotou, kvalitní potrava (což neznamená, že musí být „nadupaná“ živinami, často právě naopak), prostředí a spolubydlící minimalizující stres, v případě nutné manipulace co nejopatrnější zacházení. I kdybychom dokázali vytvořit prostředí bez patogenů, což je v běžné praxi nemožné, ještě nám tu zbývá další „nepřátelské“ médium, které naše ryby neustále „napadá“. A tím je samotná voda.
Osmóza jako spouštěč líhnutí
Osmóza má velký vliv na líhnutí – například artémie. Když dojde k namočení cysty, tak se uvnitř nastartuje metabolismus vedoucí ke vzniku glycerolu. Jeho rostoucí koncentrace uvnitř cysty vede ke změně vnitřního osmotického tlaku – pokud je rozdíl oproti osmotickému tlaku okolní slané vody značný, dojde k prasknutí obalu. Jelikož se cysty ocitají často v extrémně slaném prostředí, je nutná větší produkce glycerolu, což nauplie připraví o cenné zásoby energie. Je tedy výhodnější líhnout žábronožku při nejnižší vhodné salinitě, která ale nesmí poklesnout pod hranici tolerance vylíhnutých nauplií. Jinak proběhne sice líhnutí (resp. prasknutí obalu) v pořádku a rychle, ale obdobně rychle také nauplie v hypotonickém prostředí uhynou.
Pokud bylo smočení cysty jen přechodné a opět vyschne, než se stačila otevřít, metabolismus se zastavuje a znovu se spustí při další příležitosti. Máme-li cysty, které byly vystaveny opakovaně vlhkosti, stále ještě mohou být schopné líhnutí, ale jejich energetická zásoba je nízká. O to více je potřeba to se solí nepřehánět. Spodní hranice je podle mých osobních zkušeností zhruba 4–5 g/l, předpokládám však, že se to může lišit podle původu konkrétních vajíček.
Jiným zajímavým příkladem významu osmózy je líhnutí listonohů. I u nich vydrží cysty ve spícím stavu po několik let a čekají, až nastanou vhodné podmínky. Oproti průběhu líhnutí akvaristům důvěrně známé Artemia salina je tady ale jedna odlišnost, která vychází z podmínek přirozeného prostředí obou skupin organismů. Artémie čeká, až se ocitne ve vodě – a pokud je voda příhodné teploty, tak jí to stačí k nastartování procesů vedoucích k líhnutí. Listonoh čeká, až se ocitne ve vodě čerstvé. Vzhledem k sezónnímu a velmi krátkodobému výskytu vhodných míst signalizuje voda s nízkou salinitou čerstvé zaplavení, tedy pokyn, že teď je potřeba se vylíhnout a rychle projít celým životním cyklem. Když je voda bohatá na rozpuštěné látky, nebo dokonce salinita stoupá, cysta líhnutí nezahájí, příp. ho přeruší – lze totiž očekávat, že již dochází k odpařování vody a hrozí vyschnutí.
Toto čekání cyst na čistou vodu můžeme uplatnit v praxi, kdy k přípravě vody pro líhnutí listonohů použijeme vodu co nejměkčí, s nejnižší vodivostí. Příp. je možné vodu připravit dvoustupňově, nejprve vajíčka zalít vodou měkkou, a potom ji ještě více naředit (např. vodou destilovanou nebo z RO). Povzbudíme tím líhnutí, které využívá stejný mechanismus – produkci glycerolu, vedoucí k dostatečnému navýšení osmotického tlaku uvnitř obalu.
Podobnou „kontrolu kvality vody“, kdy salinita není jediným řídícím parametrem, významná je i teplota či množství rozpuštěného kyslíku a CO2, provádějí i další vodní organismy s diapauzou (zastavením vývoje v cystě). Existují pochopitelně i druhy žábronožek, které nejsou vázané na slanou vodu, ale naopak se líhnou v prvních jarních záplavách – např. u nás žijící žábronožka sněžná. Oba shora uvedené příklady tak demonstrují dva akvaristům nejznámější živočichy z této skupiny, ale rozhodně nepokrývají celou škálu možných nároků na vhodné podmínky.
Vliv osmózy na vývoj jiker
Jikra je krytá měkkou membránou, tzv. chorionem. Po vypuzení dochází během krátkého časového úseku (u sladkovodních ryb v řádu desítek minut) k vnikání vody dovnitř. V tomto stádiu je možné oplození. Jikra se nasátou vodou zvětší. Zároveň chorion ztvrdne, na povrchu samotného vajíčka se z cytoplazmatické membrány utváří tzv. vitelinní membrána. Prostor mezi ní a chorionem se nazývá perivitelinní prostor a je vyplněný perivitelinní tekutinou – nasátou vodou. Při prohlížení jiker pod mikroskopem nebo silnou lupou si proto můžeme všimnout, že za pár hodin po tření vlastně vidíme „kuličku v kuličce“ s jakousi čepičkou, kde je perivitelinní tekutina koncentrována. Je to obdobné jako u slepičího vejce, kde pod skořápkou najdeme další blanku, kryjící samotné vajíčko, a také dutinku mezi blankou a skořápkou.
Vnitřní, tedy vitelinní membrána se velmi rychle stává nepropustnou pro soli i vodu. V pozdějších stádiích vývoje, když už má embryo funkční vylučovací soustavu, se ale prostupnost membrány zvýší. Osmolalita vajíčka však zůstává po celou dobu vývoje prakticky stejná – a je také totožná s osmolalitou tkání dospělých ryb. Ačkoliv mechanismus osmoregulace u raných embryí ještě není plně vysvětlený, víme, že zárodek už si později (zhruba od chvíle, kdy jsou viditelné jeho oči) vyměňuje určité látky se svým okolím. Se zvyšováním propustnosti membrány dojde ke zvýšení vnitřního tlaku, a nakonec k prasknutí obalu, kdy nastává líhnutí. V extrémně hypotonickém prostředí může dojít k předčasnému prasknutí jikry, protože voda vniká dovnitř příliš intenzivně.
Pro nás je zajímavé, že proces ztvrdnutí a uzavření povrchu je rychlejší v tvrdší vodě. Pro druhy, které pocházejí z velmi měkkých vod, je proto nutné držet tvrdost (GH) co nejníže, jinak je oplozenost mizivá. Kritická je zhruba hodina po tření – poté už lze předpokládat, že jikra je oplozená, a na další vývoj už nemá tvrdost tak zásadní vliv. Každopádně se musíme vyvarovat jakýchkoliv skokových změn, které by vedly k osmotickému šoku. Nicméně odchov a samotný chov v tvrdší vodě je zpravidla dobře možný.
I když je potřeba u některých druhů snížit pro tření tvrdost blízko k nule, nějaký alespoň minimální obsah solí voda mít musí. Jinak by docházelo k poruchám vývoje jiker a/nebo jejich předčasnému prasknutí. Vždy je potřeba se řídit tím, odkud ryby pocházejí a v jakých podmínkách se po tisíciletí vyvíjely. Platí to pro dospělé ryby, které ale mají díky výkonné osmoregulaci schopnost se do značné míry přizpůsobit, byť za cenu nemalého výdeje energie, a daleko více to platí i pro jikry a embrya v raném stádiu vývoje.
Zajímavý pokus na tilápiích [1] odhalil, že zárodek si salinitu svého okolí v 2–3 hodinách po tření „zakódoval“ po zbytek svého života. Ta salinita, ve které se vyvíjel první hodiny, se ukázala jako optimální i v dospělosti a vedla k nejlepší prosperitě. Nezáleželo na tom, v jaké salinitě probíhal raný vývoj po těch úvodních hodinách, rybičky si jakoby pamatovaly výchozí hodnotu. Představuju si, že zárodek si při nasávání vody před uzavřením membrány „osahá“, kde se to vlastně narodil, a nějakým způsobem svou osmoregulaci na tuto výchozí hodnotu nastaví. Je to zajímavá adaptace na konkrétní podmínky prostředí, nicméně jsou tu dvě ALE: salinita pochopitelně musí ležet v tolerovaném rozsahu, není tudíž možné přenesením jikry sladkovodního druhu do moře rybu úplně převychovat. A adaptace je vázaná na dané individuum, není geneticky přenosná.
Ryby a tolerance salinity
Je evidentní, že salinita a osmotický tlak spolu jdou ruku v ruce – protože, jak už víme, v běžných podmínkách tvoří právě rozpuštěné soli naprostou většinu osmoticky aktivních látek. Některé ryby se dokáží přizpůsobit širšímu rozpětí salinity, ty nazýváme euryhalinní. Jejich opakem jsou ryby stenohalinní, které prosperují jen v úzkém rozpětí hodnot.
Obyvatelé moří jsou obvykle stenohalinní, ačkoliv některé druhy bezobratlých jsou v prostředí s nízkou salinitou schopní částečné osmoregulace (zatímco při plné salinitě se chovají jako ryzí osmokonformeři), takže mohou žít i v příbřežních zónách s brakickou vodou. Podobně mohou být euryhalinní i mořské ryby. Je ale nutné si uvědomit, že pokud je organismus vystavený podmínkám, které jsou na okraji jeho tolerance (tedy nejsou optimální), vynakládá na zvýšenou osmoregulaci značný díl energie a např. u produkčních ryb bylo zjištěno, že to vede k pomalejšímu růstu.
U sladkovodních ryb najdeme zástupce obou typů. Vcelku snadno si ale odvodíme, že obývá-li tetra výhradně pralesní potůčky s černou vodou, na zvýšené množství soli bude citlivá. Naopak např. cichlidy bývají považovány všeobecně za euryhalinní, protože se do sladké vody dostaly z moří relativně nedávno a mnoho druhů žije blízko ústí řek. Plně mořskou vodu nebo dokonce i slanější snáší např. ‚Cichlasoma‘ urophthalmus, Etroplus suratensis či některé druhy tilápií.
Stenohalinní druhy adaptované na měkké vody málokdy ocení přídavek soli z preventivních důvodů. Od této praxe se ustupuje i u těch skupin ryb, kde to bylo relativně běžné, jako jsou živorodky nebo africké jezerní cichlidy. Je nutné si uvědomit, že NaCl nemůže nahradit další nutné prvky (Ca, Mg, K, P, S aj.). Vzhledem k silnému vlivu NaCl na regulaci toku vody a dalších látek z/do těla ryby může přisolení pomoci ulevit od nadměrného zavodňování, jak už bylo uvedeno výše, a také snižuje toxicitu dusíkatých látek. Na opačné straně účinků je ale např. poškození vývoje zárodků při extrémním chemismu vody (nízká tvrdost). Dloouhodobé vystavení účinkům soli může vést k nedostatku jiných důležitých elektrolytů a tím paradoxně i k selhání osmoregulace a k vodnatelnosti.
Sladkomořské akvárium?!
S tímto tématem souvisí i experimenty, které se provádějí už někdy od padesátých let minulého století. Možná jste postřehli snímky z akvaristických světových veletrhů před několika lety, na nichž proplouvaly mořské a sladkovodní ryby společně v jedné expoziční nádrži. Vyvolalo to krátkodobě velkou vlnu zájmu, ale nejde o nic nového. Už kdysi se tímto směrem vydala např. americká firma Kordon, která prodávala tzv. Wonder Water. Přípravek, který měl umožnit úpravu vody právě pro spojení mořských a sladkovodních druhů. Je jasné, že obě skupiny ryb prostě ve vodě fungují úplně jinak – a jen několik výjimek, z nichž nejznámější jsou lososi, si z evidentního důvodu umějí poradit s oběma typy prostředí a umí přepnout osmoregulační mechanismy. Tady ale nešlo o amfidromní druhy, které migrují z moře do řek či naopak. Marketingovým tahákem měli být např. terčovci mezi korály. Firma Kordon na svých stránkách [2] uvádí, že ve skutečnosti tento jejich postup mohl být funkční jen pro sekundárně sladkovodní druhy, tj. takové, které se do kontinentálních vod vrátily po určitém období vývoje v moři. A jelikož to zákazníci nebyli sto pochopit a respektovat, tak vzhledem k nebývalé agresivitě mořských ryb vůči sladkovodním (která právě měla být lépe zvládána ne tak vzdáleně příbuznými, což je poněkud zvláštní argument) nakonec od prodeje tohoto produktu firma ustoupila. Z toho si můžeme odvodit, že „trvanlivější“ v tomto experimentu byly ryby mořské.
Čas od času se téma znovu otevře, když začne jiná firma zkoumat potenciál této díry na trhu. V podstatě jde o to, vytvořit takové prostředí, které nebude způsobovat potíže ani mořským rybám (nesmí být hypotonické, trpěly by dehydratací), ani sladkovodním (voda nesmí být moc slaná). Soli jsou nahrazovány organickými látkami, ponejvíce směsí sacharidů či alkoholy. Jeden patent staršího data s mnoha detaily si lze prohlédnout tady [3].
Nedá mi to nevyjádřit zde můj ryze subjektivní názor, že snaha o obalamutění přírody za účelem vytvoření něčeho „jedinečného“ a výstředního nemá daleko k výrobě ryb tetovaných, barvených a jinak fyzicky mrzačených. Zatímco v prvním případě nebyl marketingový tah příliš úspěšný a ryby spolupracovat nechtěly – pohled na utrápenou mečovku, jak se houpe v rohu nádrže a marně se brání útokům králíčkovce, než to za několik týdnů vzdá, nepřipadal cool ani těm nejotrlejším majitelům – srdíčkové a tyrkysové a růžové ryby jsou tak roztomilé… a jdou tak na dračku.
Ve skutečnosti máme ještě obrovské mezery v našich znalostech toho, jak ryby žijí a jak je máme chovat. Poznávat jejich přirozené prostředí a potřeby a obdivovat jejich krásu, pokud se dobře cítí, je rozhodně naplňujícím koníčkem.
Reference: [1] Watanabe, W.O., Kuo, C.-M. & Huang, M.-C. (1984): Experimental rearing of Nile tilapia fry (Oreochromis niloticus) for saltwater culture. ICLARM, p. 25. [2] www.kordon.com/kordon/news/wonder-water [3] www.freepatentsonline.com/3683855.html [4] Oldfield, R.G. (2004): Salty cichlids. Knowledge of salinity tolerance and preference may allow new species combinations and improved husbandry in aquaria. Freshwater and Marine Aquarium. 27(8): 98-104. [5] Krogh, A. (2015): Osmotic Regulation in Aquatic Animals. Reprint from 1939. Cambridge University Press, 252 pp. [6] McNab, B.K. (2002): The Physiological Ecology of Vertebrates: A View from Energetics. Cornell University Press, 576 pp. [7] Evans, D.H. & Claiborne, J.B. (2005): The Physiology of Fishes, 3rd edition. CRC Press, 616 pp.© 2016

